Scientists Predict Earth-Like Habitable Exoplanet Will Be Found in 2011
An artist’s impression of Gliese 581d, an exoplanet about 20.3 light-years away from Earth, in the constellation Libra. Credit: NASA
[/caption]
Two astronomers have written a paper and say that the first Earth-like, habitable exoplanet will be announced in May of 2011. Do they have inside information, a crystal ball, or amazing powers of prediction? No, they base their projection on math and trends from the past 15 years of exoplanet discoveries. And if the discoveries continue at their present rate, the researchers say next year is the year of the long awaited holy grail of finding another Earth-like planet out in the cosmos.
Samuel Arbesman from Harvard Medical School in Boston and Gregory Laughlin at the University of California, Santa Cruz take a scientometric approach to their prediction. Scientometrics is the science of measuring and analyzing science, and is often done using bibliometrics which is a measurement of the impact of scientific publications. Arbesman and Laughlin said this type of work highlights the usefulness of predictive scientometric techniques to understand the pace of scientific discovery in many fields.
They use the properties of previously discovered exoplanets along with external estimates for the discovery of the first potentially habitable extrasolar planet.
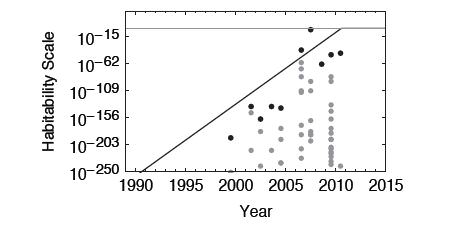
In their paper they indicate that since astronomers have been discovering extrasolar planets at an increasing rate since 1995 and the discoveries follow a well understood pattern, it should be easy to predict when planet searchers will hit the jackpot.
The first exoplanets found were the massive Jupiter or larger-sized planets which were the easiest to find, and then as techniques improved over the past 15 years, astronomers have found smaller planets, some just a few times more massive than Earth.
Arbesman and Laughlin took that rate of discovery, and they also needed to factor in all the variables for what we think will make a planet habitable: the surface temperature must allow liquid water to exist, so that life as we know it can appear, and that depends on the size of the star, how far the planet orbits from its star, and what type of surface the exoplanet has.
They conclude there is a 66 per cent probability of finding another Earth by 2013, a 75 per cent probability by 2020, and a 95 per cent probability by 2264, but the median date of discovery is in May 2011. And not just sometime in May, but “early May.”
In June 2010, the Kepler Telescope team revealed they had found 750 exoplanet candidates, and a fair number of those confirmed might be Earth-sized. They expect they can confirm and announce some of these candidates in February 2011. But Arbesman and Laughlin predict it might take longer. “Because of the limited time base line of the mission to date, the Kepler planet candidates to published in February 2011 may be too hot to support significant values for H (which is their habitability metric),” they wrote in their paper.
So, if their prediction comes true, that might mean another team, such as the HARPS, or Keck, or CoRoT, or other exoplanet-finding wizards might make the discovery.
“It must be noted that by publicizing our prediction, there is a concern that it will become accurate,” Arbesman and Laughlin write in their paper, “simply due to the well-studied Hawthorne Effect. However, due to the large number of observations and long periods of time required to confirm an extrasolar planet discovery, it is unlikely that our prediction at this time will appreciably affect the announcement of the discovery of an Earth-like planet. Therefore, it is reasonable to use the habitability metric curve as a rough prediction for when the first potentially habitable planet will be discovered, in this case, as early as May 2011, and likely by the end of 2013.”
It will be interesting to see how accurate their prediction turns out to be!
Additional Source: Technology Review Blog
Recent Posts
Update your Desktop Wallpaper with 25 New Images from Chandra
It’s not always possible to observe the night sky from the surface of the Earth.…
SpaceX Resumes Falcon 9 Rocket Launches After FAA Go-Ahead
SpaceX is flying again after the Federal Aviation Administration ruled that the company can resume…

{kind=link}
Is This How You Get Hot Jupiters?
When we think of Jupiter-type planets, we usually picture massive cloud-covered worlds orbiting far from…
Now Uranus’ Moon Ariel Might Have an Ocean too
Venus is known for being really quite inhospitable with high surface temperatures and Mars is…
Why is JWST Having So Much Trouble with the TRAPPIST-1 System?
When the James Webb Space Telescope was launched it came with a fanfare expecting amazing…
Planetary Habitability Depends on its Star’s Magnetic Field
The extrasolar planet census recently passed a major milestone, with 5500 confirmed candidates in 4,243…